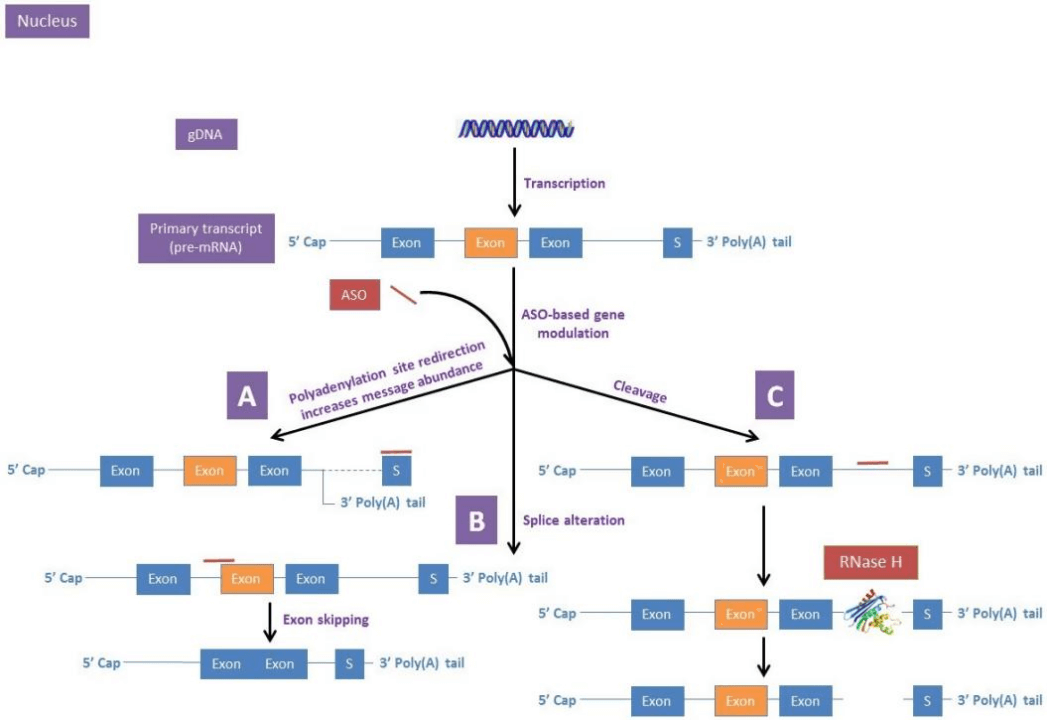
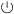Service














Gene Synthesis Services
Oligonucleotide Synthesis Services
Sanger Sequencing Services
Recombinant Protein Expression Service
Recombinant Antibody Expression Service
Pharmaceutical NGS Safety Testing Services
NGS Research and Technology Services
Exome Sequencing
Whole Human Genome Resequencing
Whole Genome Resequencing Of Plants And Animals
Eukaryotic/prokaryotic Transcriptome Sequencing
Small RNA Sequencing
Long non-coding RNA (lncRNA sequencing)
Full-length 16s, 18s, and ITS Amplicon Sequencing
Metagenomic Sequencing
Macro-genome Sequencing
Bacterial/Fungal WGS Sequencing
Bacterial/Fungal De novo Sequencing





















EN